近日,永利hb99402菊花遗传与种质创新团队的蒋甲福教授在国际知名期刊The Plant Journal发表了题为“The RAV transcription factor TEMPRANILLO1 involved in ethylene-mediated delay of Chrysanthemum flowering”的论文,揭示了菊花CmTEM1介导乙烯抑制菊花开花的分子调节机制。
“花开不并百花丛,独立疏篱趣未穷” 。菊花(Chrysanthemum morifoliumRamat.)是世界范围广泛栽培和应用的观赏植物,多数菊花品种为短日照植物,开花时间多集中于秋季。目前,虽可通过光温调节进行花期调控以实现菊花周年生产,但反季节生产的高能耗成本严重制约了菊花产业的健康发展。在生产中常用适当浓度的乙烯利辅助加光处理以推迟菊花开花,但其调控的分子机制并不清楚。

RELATED TO ABI3 AND VP1(RAV)转录因子家族是植物特有的一类转录因子,参与调控多个发育和生理过程。TEMPRANILLO 1(TEM1)是RAV转录因子家族的一员,具有转录抑制活性,参与多种开花途径,抑制幼年期向成年期转变。本研究从菊花中分离并鉴定了AtTEM1的同源基因,命名为CmTEM1,研究表明CmTEM1受乙烯诱导上调表达;相较于清水处理的对照,经外源乙烯利处理的野生型菊花晚花11.1 d,两个沉默株系amiR-CmTEM1分别晚花2.1 d和5.2 d,而两个超表达株系CmTEM1-OX分别晚花23.9 d和28.1 d(图1)。这表明沉默株系amiR-CmTEM1对乙烯的敏感性降低,而超表达株系CmTEM1-OX对乙烯更敏感,说明CmTEM1参与乙烯调控菊花开花。
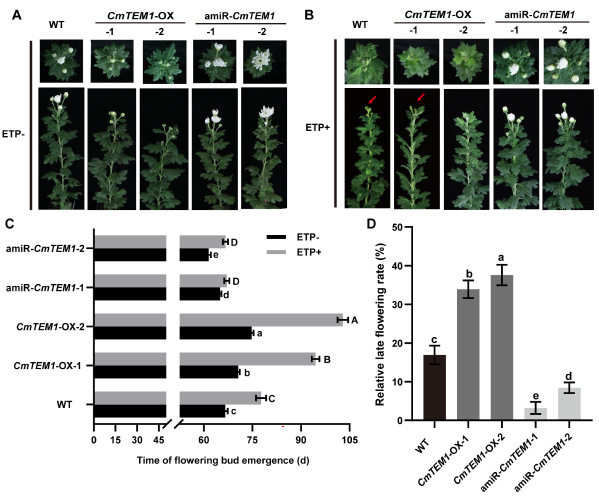
图1 乙烯处理野生型和CmTEM1转基因菊花的开花表型
进一步对超表达株系CmTEM1-OX进行转录组测序分析,发现作为APETALA1(AP1)/FRUITFULL(FUL)亚家族成员的开花整合子CmAFL1的表达显著下调,RT-qPCR实验证实该基因在沉默株系amiR-CmTEM1中上调表达。分子生物学实验表明CmTEM1能直接结合CmAFL1的启动子并抑制其转录。为了进一步明确CmTEM1与CmAFL1之间的调控关系,利用甘蓝卷叶病毒沉默载体表达系统分别在WT和amiR-CmTEM1株系中沉默CmAFL1,表型观察发现CaLCuV-CmAFL1/ amiR-CmTEM1植株开花早于CaLCuV-CmAFL1/WT植株,但晚花于CaLCuV/WT植株(图2)。
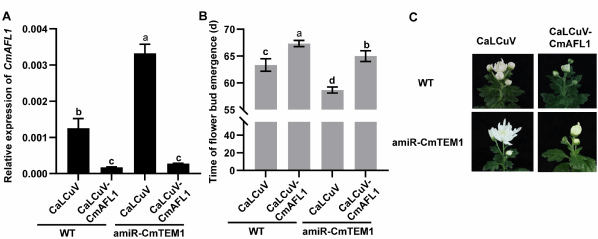
图2 CaLCuV-CmAFL1转化野生型和amiR-CmTEM1沉默菊花株系的表型
综上所述,该研究揭示了CmTEM1部分依赖CmAFL1介导乙烯抑制菊花开花的分子机制(图3)。研究结果为深入理解乙烯抑制菊花开花提供了新的认知,为菊花周年生产提供了理论指导。
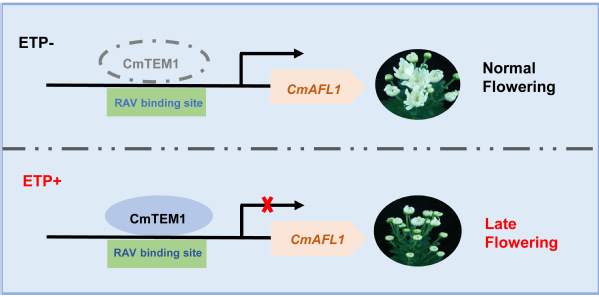
图3 CmTEM1-CmAFL1介导乙烯抑制菊花开花的工作模式图
永利hb99402陈发棣教授领衔的“菊花遗传与种质创新团队”围绕开花优异基因挖掘与分子改良开展了较为深入的创新性研究。发现夏菊花期调控机制与秋菊不同,CmFTL1是夏菊在长日照条件下的开花素,且被CmBBX8直接转录调控,推测该BBX8-FT模块在夏菊开花调控中发挥重要作用(Plant Biotechnology Journal 2020);CmBBX8 协同CmRCD1和CmERF3 精准调控CmFTL1的表达,防止夏菊从营养生长向生殖生长转换过快,而导致切花品质不良(Horticulture Research 2021,Plant Cell and Environment 2022)。发现秋菊中CmERF110和CmFLK共同通过响应光周期和调控生物钟变化促进菊花开花(Plant Cell and Environment 2022);而CCT家族蛋白CmNRRa与14-3-3家族蛋白Cm14-3-3μ协同抑制光周期路径(Journal of Experimental Botany 2023);CmSVP招募CmTPL抑制短日照下开花素基因CmFTL3的表达而调控秋菊低温下的开花(Plant Physiology 2023)。本论文进一步揭示了光周期途径中,菊花受乙烯抑制开花的调控机制,为培育节能菊花新品种提供了优异基因资源。
永利hb99402蒋甲福教授为该论文的通讯作者,永利hb99402已毕业博士生程华为论文第一作者,菊花遗传与种质创新团队的周李杰讲师、宋爱萍副教授,王利凯教授、陈素梅教授和陈发棣教授等参与了研究工作。该研究得到了国家自然科学基金(31930100、32272756) 等项目的资助。

